
Fantastic news Tom, no doubt they are stoked to have you!
16.04.2025 15:46
👍 1
🔁 0
💬 0
📌 0
Fantastic news Tom, no doubt they are stoked to have you!
BOLD variability (especially on-task) typically has loads of behaviourally-relevant effects, at within- and between-subject levels. No amount of preprocessing/noise removal can kill those effects in our hands. For example: www.cell.com/neuron/pdf/S...
The majority of variance in BOLD has nothing to do with motion. MeanBOLD and SDBOLD are rarely highly correlated in any dataset I've seen, any ratio of the two (e.g., tSNR) makes interpretations unnecessarily difficult. Could start by linking mean and SD estimates you have separately to behaviour?
This is happening tomorrow! 😅😅😅
Agreed...No doubt film is a major step forward over resting state, and is indeed an engaging task in its own right, especially when behavior is concurrently collected!
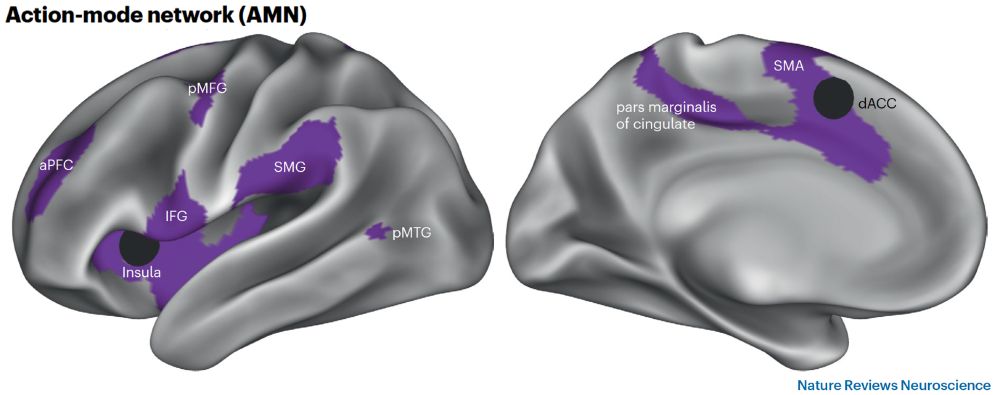
The brain’s action-mode is created by a dedicated action-mode network (AMN) rdcu.be/d5odm. In the brain’s mode continuum, AMN sits opposite DMN’s default-mode, as yin-yang. AMN might be key in pain, apathy, Parkinson’s. New @natrevneurosci.bsky.social w/ Marc Raichle & @gordonneuro.bsky.social 🧵 ⬇️

Thanks a lot Ellie! Film might be logistically easier, but clinical meaningful, disorder-relevant tasks are grossly underutilised in our opinion. Even incredibly short tasks can be highly effective and relatively easy to implement...for example: www.biologicalpsychiatryjournal.com/article/S000...

To start off 2025, we are very pleased to introduce a Special Issue on the Neuroscience of Aging, curated by Axel Guskjolen.
We hope you enjoy every single piece of it as much as we do!
www.cell.com/neuron/current
PhD student @zoyamooraj.bsky.social et al. with a remarkable vision for the future of the cog neuro of human aging, now live in Neuron: www.cell.com/neuron/fullt....
Task-related function is the future y'all... there can be no substitutes if we want to understand cognitive aging.
Curious to hear your take on the structure-function stance as well...no doubt you're one of those trying to do it right. Keep up the great work guy😤
Outstanding man, congrats!
New LIP paper out now: Congratulations to colleagues Leo Waschke and Fabian Kamp and co-authors!
Massive thanks to all co-authors (Waschke, Kamp, van den Elzen, Krishna, Lindenberger, and Rutishauser) and to @lipmpib.bsky.social @mpib-berlin.bsky.social for support of one of the most important projects I have had the pleasure to work on. Cheers!

We thus propose that moment-to-moment spiking variability may provide a new window into how the hippocampus constructs memories from the "building blocks" of our sensory world.

Crucially, we show that the more precisely hippocampal spiking variability tracks the composite (late-layer) features that comprise each individual stimulus, the better those stimuli are later remembered up to 30 minutes later. These effects were also *spatially specific* to the hippocampus.

Hippocampus spiking entropy was coupled to image features in every.single.subject...with tighter coupling to more composite (late layer) than to simpler (early layer) image features.

We used comp. vision models (HMAX & VGG16) to estimate the features of images presented to patients (N = 34) during memory encoding. We then estimated the coupling of hippocampal single-neuron variability (entropy) to image features via within-participant latent correlations.

Ok, but what signature of hipp. spiking might track such features? We've shown that the variability of fMRI activity in visual cortex scales with the complexity of visual input (sciencedirect.com/science/arti...). Could this also be the case in the hippocampus during memory formation?

In turn, this permits direct testing of the “sensory feature space” the hippocampus leverages during memory formation.

We argue that the architecture of multi-layer computational vision models can be used to differentiate between simple and composite visual features of any stimulus a participant may encode.
More composite features might dominate due to the hippocampus’ hierarchical position and afferent projections, but “simple” features might still be crucial for detailed representations. How can we know?

During memory formation, the hippocampus is presumed to represent and conjunct the “content” of stimuli. How does it do this? On which kind of sensory features does the hippocampus rely when memories are formed?
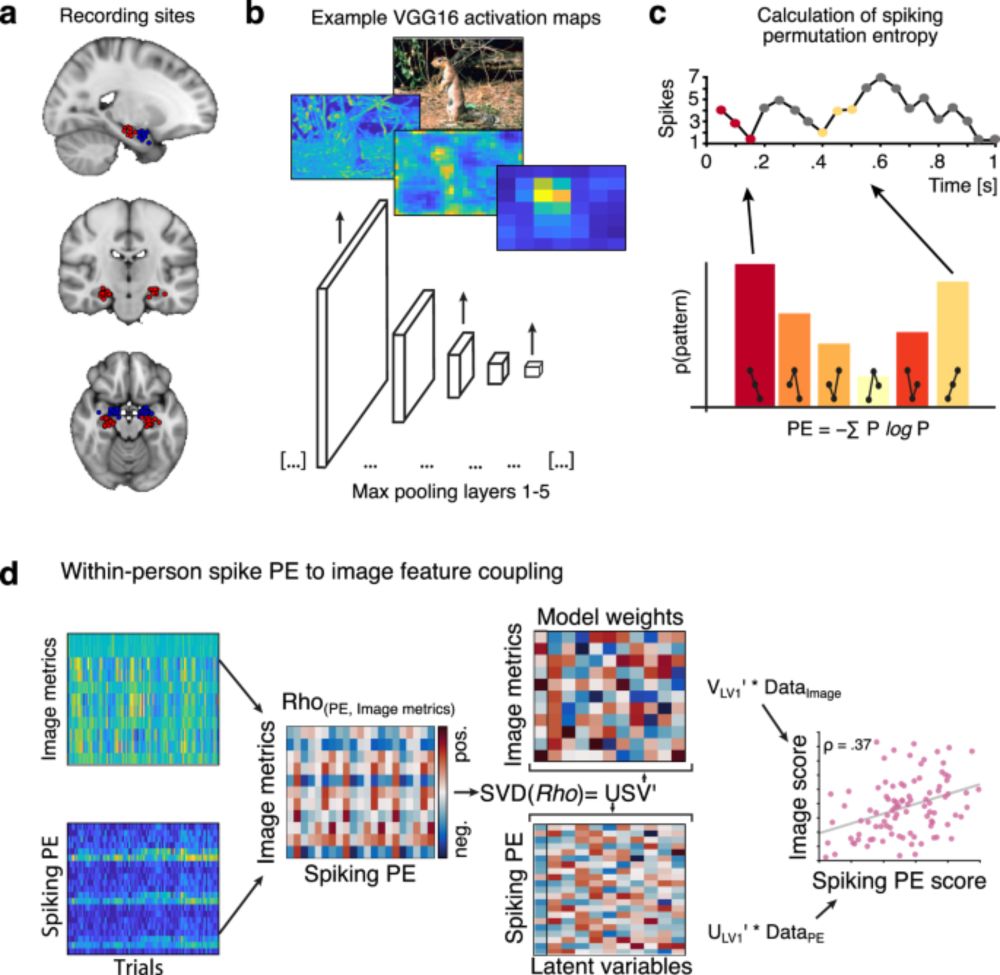
*How* does the hippocampus encode the world around us? Leo Waschke and Fabian Kamp bring the 🔥, showing that single-neuron spiking variability in hippocampus dynamically tracks sensory content during memory formation in humans. Our latest in Nature Communications: www.nature.com/articles/s41...
JEALOUS. Enjoy!
Absolutely outstanding multi-modal work on uncertainty in the aging brain by @juliankosciessa.bsky.social et al., many years in the making. EEG, fMRI, comp modelling, sub vs. cortical, this one has it all!
Now live in Nature Communications: www.nature.com/articles/s41...